Related posts:Highlights
• Sexual selection has been stronger in humans than is often assumed.
• Ancestral men competed primarily through force and threat of force.
• Ancestral women competed mainly through mate attraction.
• Understanding sexual selection clarifies some human psychological sex differences.Abstract
Sexual selection favors traits that aid in competition over mates. Widespread monogamous mating, biparental care, moderate body size sexual dimorphism, and low canine tooth dimorphism suggest modest sexual selection operating over human evolution, but other evidence indicates that sexual selection has actually been comparatively strong. Ancestral men probably competed for mates mainly by excluding competitors by force or threat, and women likely competed primarily by attracting mates. These and other forms of sexual selection shaped human anatomy and psychology, including some psychological sex differences.
"Ancestral men probably competed for mates mainly by excluding competitors by force or threat"
Physical attractiveness as a phenotypic marker of health
Evolutionary explanations regarding the differential preference for particular traits hold that preferences arose due to traits’ association with increased potential for reproductive fitness. Assessments of physical attractiveness have been shown to be related to perceived and measured levels of health, an important fitness-related trait. Despite the robust association between physical attractiveness and health observed in the extant literature, a number of theoretical and methodological concerns remain. Specifically, the research in this area possesses a lack of specificity in terms of measures of health, a reliance on artificial social interactions in assessing physical attractiveness, a relatively infrequent use of non-student samples, and has left unaddressed the confounding effects of raters of attractiveness. Using these concerns as a springboard, the current study employed data from the National Longitudinal Study for Adolescent Health (N ≈ 15,000; aged 25 to 34 years) to assess the relationship between physical attractiveness and various specific and overall measures of health. Logistic and OLS regression models illustrated a robust association between physical attractiveness and various measures of health, controlling for a variety of confounding factors. In sum, the more attractive a respondent was rated, the less likely he or she was to report being diagnosed with a wide range of chronic diseases and neuropsychological disorders. Importantly, this finding was observed for both sexes. These analyses provide further support for physical attractiveness as a phenotypic marker of health. The findings are discussed in reference to evolutionary theory and the limitations of the study and future research suggestions are also addressed.
"Monkeys' faces evolved to avoid crossbreeding"
Dr James Higham, senior author, said: "Evolution produces adaptations that help animals thrive in a particular environment, and over time these adaptations lead to the evolution of new species."A key question is what mechanisms keep closely related species that overlap geographically from interbreeding, so that they are maintained as separate species.
"Our findings offer evidence for the use of visual signals to help ensure species recognition: species may evolve to look distinct specifically from the other species they are at risk of interbreeding with," Dr Higham said.
"In other words, how you end up looking is a function of how those around you look. With the primates we studied, this has a purpose: to strengthen reproductive isolation between populations."
SMBE 2014: Insights on Sexually Antagonistic Selection in the Human Genome
1Eco-anthropologie et Ethnobiologie, UMR 7206 CNRS, MNHN, Univ Paris Diderot, Sorbonne Paris Cité, F-75005, Paris, France, 2Max Planck Institute for Evolutionary Anthropology, Department of Evolutionary Genetics, Leipzig, Germany
ESEB 2013 abstracts and videos
Genetic genealogy comes of age: advances in the use of deep-rooted pedigrees in human evolutionary research (video)
Author(s): Larmuseau, MHD, Van Geystelen, A, Decorte, R
Summary:
Research on the recent human evolution will benefit from the implementation of extended genetic genealogical data. The approach to combine deep-rooted pedigrees with genetic information advances the understanding of changes in the human population genetic structure during the last centuries. This recent advance is mainly based on the extensive growth of whole genome sequencing data and available genealogical data of high quality. Moreover, according to the latest genetic genealogical research the historical non-paternity rate in Western Europe is estimated around 1% per generation within the last four centuries, which means that the expected relationship between the legal genealogy and the genetics of DNA donors exists. Therefore, genetic genealogical data will help with three research aims of human evolutionary studies: (I) detecting signals of (past) population stratification and interpreting the population structure in a more objective manner, (II) obtaining the time scale and impact of particular detected gene flow events more accurately and (III) determining temporal genetic differentiation within a population by combining in-depth pedigree data with haploid markers. Each of these research aims will be discussed with examples of the human population in Flanders (Western Europe). At the end, we will discuss the advantages and pitfalls of using genetic genealogy within studies on human evolutionary genomics.
Detection of polygenic selection at different evolutionary levels (video)
Author(s): Excoffier L, Daub J
Summary:
Most approaches aiming at finding genes involved in adaptive events have focused on the detection of outlier loci, which resulted in the discovery of individually ´significant´ genes with strong effects. However, a collection of small effect mutations could have a large effect on a given biological pathway that includes many genes, and such a polygenic mode of adaptation has not been systematically investigated in humans or other mammals. We therefore propose to evidence polygenic selection by detecting signals of adaptation at the pathway or gene set level instead of analyzing single independent genes. Using a gene-set enrichment test, we identify genome-wide signals of recent adaptation among human populations as well as more ancient signals of adaptation in the human lineage and in primates.
A genome-wide scan for relaxation of constraints in the human lineage affecting specific functional processes (video)
Author(s): Somel, M, Wilson-Sayres, M, Jordan, G, Huerta-Sanchez, E, Fumagalli, M, Ferrer-Admetlla, A, Nielsen, R
Summary:
Changes in the subsistence mode of a species can lead to adaptive evolution of new functions, while it can also cause relaxed negative selection in previously essential functions. While positive selection in humans has been intensely studied, functional processes subject to relaxed constraints in the human lineage remain largely unknown. Here we present a framework for detecting relaxation of selective constraints that affect a particular functional process specifically in one taxon. Jointly using human and chimpanzee population genomic data with mammalian comparative genomic data, we identify olfactory receptors and proteasome subunits as candidates of relaxed constraints in humans: both gene sets contain high frequency non-synonymous mutations in humans while having conserved amino-acid sequences across other mammals. We further discuss the possible underlying causes of this signal.
Selection on penis size, body shape and height in humans: a simple multivariate method to quantify female preferences based on male physical attractiveness (video)
Author(s): Mautz, BS, Jennions, MD, Peters, RA, Wong, BBM
Summary:
Compelling evidence from many animal taxa indicates that male genitalia are often under post-copulatory sexual selection for characteristics that increase a male’s relative fertilization success under sperm competition. There could, however, also be direct pre-copulatory female mate choice based on male genital traits. Before clothing, the non-retractable human penis would have been conspicuous to potential mates. This, in combination with claims that humans have a large penis for their body size compared to other primates, has generated suggestions that human penis size partly evolved due to female choice. We presented women with digitally projected fully life-size, computer-generated animations of male figures to quantify the (interactive) effects of penis size, body shape and height on female assessment of male sexual attractiveness. We generated 343 male figures that each had one of seven possible values for each of the three test traits (7x7x7 = 343). All seven test values per trait were within two standard deviations of the mean based on a representative sample of males. We calculate response (fitness) surfaces based on the average attractiveness rank each of the 343 male figure received. We also calculated individual response surfaces for 105 women (each women viewed 53 figures). Both methods yielded almost identical results. We discuss our finding in the context of previous studies that have taken a univariate approach to quantify female preferences. We discuss the hypothesis that pre-copulatory sexual selection might play a role in the evolution of genital traits.
Quantitative genetic variation, selection and secular change of skull shape in humans
Author(s): Klingenberg, C, Martínez-Abadías, N, Esparza, M, Sjøvold, T, Hernández, M
Summary:
The combined use of geometric morphometrics and quantitative genetics provides a set of powerful tools for obtaining quantitative information that is crucial for many important questions concerning the evolution of shape. In particular, the demographic information that is available for human populations make humans a unique study system for studying the mechanisms of evolutionary change in morphological traits. We investigate skull shape in the population of Hallstatt (Austria), where a collection of human skulls with associated records offer a unique opportunity for such studies. We use an individual-based statistical model to estimate the genetic covariance matrix, and characterize selection using fitness estimates from demographic data. We find clear evidence for directional selection, but not for nonlinear selection (stabilizing or disruptive selection). The predicted response to this selection, computed with genetic parameters from the population, does not match the estimate of secular change over the 150-year range of the data. We discuss possible reasons for the mismatch.
Heritability of facial attractiveness and masculinity-femininity
Human facial attractiveness and facial sexual dimorphism (masculinity–femininity) are important facets of mate choice and are hypothesized to honestly advertise genetic quality. However, it is unclear whether genes influencing facial attractiveness and masculinity–femininity have similar, opposing, or independent effects across sex, and the heritability of these phenotypes is poorly characterized. To investigate these issues, we assessed facial attractiveness and facial masculinity–femininity in the largest genetically informative sample (n = 1,580 same- and opposite-sex twin pairs and siblings) to assess these questions to date. The heritability was ~0.50–0.70 for attractiveness and ~0.40–0.50 for facial masculinity–femininity, indicating that, despite ostensible selection on genes influencing these traits, substantial genetic variation persists in both. Importantly, we found evidence for intralocus sexual conflict, whereby alleles that increase masculinity in males have the same effect in females. Additionally, genetic influences on attractiveness were shared across the sexes, suggesting that attractive fathers tend to have attractive daughters and attractive mothers tend to have attractive sons.
Individual identity signaling in humans
Michael Sheehan Morphological and population genomic evidence of selection for individual identity signaling in human faces
There's no abstract, but one area where I suspect selection of this sort may turn out to be relevant (at least more relevant than Peter Frost-style sexual selection) is in explaining European hair and eye color variation.
"Traits signaling identity should be highly variable, often display polymodal distributions, not be condition dependent (i.e., be cheap to produce and/or maintain), not be associated with fitness differences, exhibit independent assortment of component characters, and often occur as fixed phenotypes with a high degree of genetic determination.""Is human facial distinctiveness an adaptive signal of individual identity? From a sociobiological perspective, humans seem to have the ‘perfect storm’ of selection pressures that might favor recognizability. We are extremely social, interacting repeatedly with large numbers of individuals, each with varying roles in our lives. We are extremely cooperative, and we make complex decisions about whether and how much to cooperate based on kinship, friendship and social reputation [39,78]."
Self-resemblance and attractiveness
Opposing forces influence assortative mating so that one seeks a similar mate while at the same time avoiding inbreeding with close relatives. Thus, mate choice may be a balancing of phenotypic similarity and dissimilarity between partners. In the present study, we assessed the role of resemblance to Self’s facial traits in judgments of physical attractiveness. Participants chose the most attractive face image of their romantic partner among several variants, where the faces were morphed so as to include only 22% of another face. Participants distinctly preferred a “Self-based morph” (i.e., their partner’s face with a small amount of Self’s face blended into it) to other morphed images. The Self-based morph was also preferred to the morph of their partner’s face blended with the partner’s same-sex “prototype”, although the latter face was (“objectively”) judged more attractive by other individuals. When ranking morphs differing in level of amalgamation (i.e., 11% vs. 22% vs. 33%) of another face, the 22% was chosen consistently as the preferred morph and, in particular, when Self was blended in the partner’s face. A forced-choice signal-detection paradigm showed that the effect of self-resemblance operated at an unconscious level, since the same participants were unable to detect the presence of their own faces in the above morphs. We concluded that individuals, if given the opportunity, seek to promote “positive assortment” for Self’s phenotype, especially when the level of similarity approaches an optimal point that is similar to Self without causing a conscious acknowledgment of the similarity. [. . .]Current psychological research on human attractiveness has replaced the relativistic belief that “beauty is in the eye of the beholder” with a universalistic one. [. . .]
However, the opposition between the relativistic and the universalistic perspectives may only be apparent, since one can posit the coexistence of an early, developmental, “imprinting” for physical traits of close con-specifics (typically, family members but also Self) as another universal mechanism that accounts for kin recognition as well as having an impact on mating preferences [5]. Indeed, face recognition mechanisms are heritable [6] and humans may be born with a schematic knowledge of the human face, which is then modified or filled out through exposure to human faces early in life. Thus, on one hand, a facial attribute like averageness would be based on a lifetime exposure to a large number of other con-specifics [7], so that one would expect that individuals within the same social group would tend to share a very similar (or seemingly “universal”) sense of what is the human average appearance. On the other hand, an imprinting mechanism, based on early experience, would lead to the opposite effect of establishing idiosyncratic “ideals” of beauty that may differ considerably between individuals. Thus, the coexistence of general learning mechanisms and mechanisms of kin recognition should shape ideals of facial (or bodily) aesthetics that are to a great deal consistent across many individuals but contain some elements that are unique to each individual.
The beautiful woman in medieval Iberia: Rhetoric, cosmetics, and evolution
Literary portraits of the beautiful woman in medieval Iberia tend to emphasize several physical features, such as long, blond hair, or light-colored and hairless skin. This study examines the specific features of the beautiful woman in several major works and genres from medieval Iberia. It also traces the rhetorical sources of these portraits to the Classical and medieval Latin traditions, whose influence is evident in other early vernacular literatures of Europe. It then analyzes several medieval cosmetic treatises in Latin and in vernacular languages that attest to medieval women's beautifying practices, such as the use of hair-dyes, depilatories, and skin-whitening creams.
The comparison of the literary and cosmetic evidence shows a canonical view of feminine beauty that encompasses different cultural areas in medieval Iberia. This view is also consistent with ancient as well as with twenty-first century conceptions of beauty. The findings suggest that the ideal of feminine beauty in medieval Iberia is not unique, but rather a manifestation of near-universal male preferences shaped by sexual selection in the course of human evolution. [. . .]
Most cosmetic treatises devote considerable space to the maintenance of well-groomed, long, and healthy hair. They also include many recipes for hair-dyes (blond and black). In Spanish literature, blond hair appears to be more typical of learned poetry and appears to be associated with nobility: "rruvios, largos cabellos / segund doncellas d’estado" (Marqués de Santillana 11-12). In the cantigas it is not mentioned, and in the Andalusian and Arabic tradition hair is black, not without exceptions (see Chapter Three).
[Claudio Da Soller. The beautiful woman in medieval Iberia: Rhetoric, cosmetics, and evolution. University of Missouri - Columbia, 2005.]
Light-skinned black women rated more attractive

Row H3IR1 S35Q1 PHYSICAL ATTRACTIVENESS OF R-W3
Column H3IR17 S35Q17 RESPONDENT SKIN COLOR-W3
Control BIO_SEX3 BIOLOGICAL SEX-W3
Filter H3OD4B(1) S1Q4B RACE-BLACK/AFRICAN AM-W3(=Marked)
Please make no unkind inferences concerning a possible relationship between degree of European admixture and attractiveness. The obvious conclusion is Kanazawa's critics were right: biased raters downgrade the beauty of African American women owing to their racism and brainwashed adherence to Eurocentric beauty standards. This insidious form of racism apparently extends to more literal sexual marketplaces:
Gwen and Alicia were especially coveted because of their skin color. In a rigid hierarchy that clinical psychologist Melissa Farley—founder of Prostitution Research & Education, a San Francisco–based think tank—calls “eroticized racism,” the “snow bunnies” (white girls) outclass the “ducks” (black girls). “Maybe one out of 50 callers would request a black or Latina,” says Caroline. “Most asked for ‘the girl next door’—a blonde, thin teenager with big breasts. That’s candy to ants.” [Sex Trafficking of Americans: The Girls Next Door]Note: Kanazawa's methods may well be flawed. I personally wouldn't be comfortable drawing the conclusion he did based on the data he did. I actually expect racial differences (or similarities) in average rated physical attractiveness have little to do with actual sexual attraction/behavior. To the extent that ratings of physical attractiveness reflect subconscious judgments of youth, health, symmetry, and so on, we should be able to meaningfully rank attractiveness across races; but this does not make even an "attractive" black woman an ideal or preferred sexual partner for a normal white man. Sorry Severn (commenter at Mangan's who is flipping out about Kanazawa's "bad science" and who coincidentally says he has dated black women).
I'll sheepishly admit I'm a longtime regular at strip clubs. I'll often get black strippers with slightly desperate looks on their faces sitting at my table trying really hard to be friendly to me hoping they can sell me a lap dance. Meanwhile the blonde strippers in the club don't even have to approach anyone; the male customers swarm the stage when they get up to dance. In a strip club it's kind of obvious who men find the most attractive. I admit this is not a scientific study, just my personal observation. [A comment at Mangan's]
Black women rated less physically attractive than other women; white men rated better looking than black men
Kanazawa explains:
The physical attractiveness of each Add Health respondent is [rated] three times by three different interviewers over seven years.
From these three scores, I can compute the latent "physical attractiveness factor" by a statistical procedure called factor analysis. Factor analysis has the added advantage of eliminating all random measurement errors that are inherent in any scientific measurement. The latent physical attractiveness factor has a mean of 0 and a standard deviation of 1.
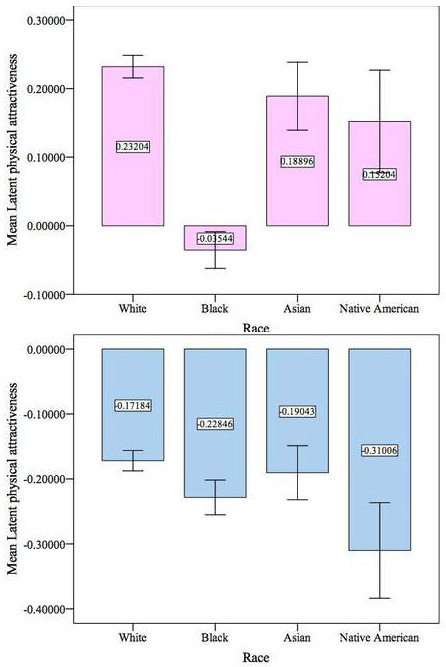
Black women are still less physically attractive than nonblack women net of BMI and intelligence. Net of intelligence, black men are significantly more physically attractive than nonblack men.Inconveniently for black men and Kanazawa, the real world does not control for intelligence. Further, if the association between intelligence and physical attractiveness rests on some underlying "good genes" factor, it makes little sense to try to "control" for intelligence when comparing races. The intelligence and attractiveness of someone with average genes for his race are what they are.
Blonde women earn more, have husbands who earn more
Our data source is the 1979 cohort of the National Longitudinal Survey of Youth (NLSY79). [. . .] We limit our sample to Caucasian women aged 25 and over. [. . .] Regression results indicate that blonde women receive a wage premium equivalent in size to the return for an extra year of schooling. A significant blondeness effect is also evident in the marriage market. Blonde women are no more or less likely to be married; but, their spouse's wages are around 6% higher than the wages of other spouse's.
Google ngrams





Masculinity, skin color, and male facial attractiveness
The proposal [. . .] that masculine men are immunocompetent and attractive – underpins a large literature on facial masculinity preferences. Recently, theoretical models have suggested that current condition may be a better index of mate value than past immunocompetence. This is particularly likely in populations where pathogenic fluctuation is fast relative to host life history. As life history is slow in humans, there is reason to expect that, among humans, condition-dependent traits might contribute more to attractiveness than relatively stable traits such as masculinity. [. . .]The authors point out problems with attempts to assess the affect of masculinity on facial attractiveness that rely on human ratings of perceived masculinity or digital manipulation of photographs: (1) for rated masculinity, "subjective judgments of masculinity are based on factors other than just morphological masculinity"; (2) with morphing techniques, factors potentially more important than masculinity in determining real world attractiveness are not allowed to vary, and a preference for averageness might result in participants systematically preferring more or less masculine morphs even if women are completely indifferent to masculinity. As for the effects of skin color, in this sample:
The relationship between masculinity and attractiveness was assessed in two samples of male faces. Most previous research has assessed masculinity either with subjective ratings or with simple anatomical measures. Here, we used geometric morphometric techniques to assess facial masculinity, generating a morphological masculinity measure based on a discriminant function that correctly classified >96% faces as male or female. When assessed using this measure, there was no relationship between morphological masculinity and rated attractiveness. In contrast, skin colour – a fluctuating, condition-dependent cue – was a significant predictor of attractiveness.
The regression retained only skin yellowness as a predictor of attractiveness, and the effect of skin yellowness was positive and highly significant (F(1,71) = 10.806, Beta = .366, t = 3.287, p<.002). Skin lightness, redness and morphological masculinity did not significantly predict attractiveness (all p>.114, see Table 1).Other studies have also found increased skin lightness and redness associated with perceived health and attractiveness. The association of yellowness with attractiveness "may be attributable to dietary carotenoid deposition in the skin. This suggests that carotenoids, which are involved in health signaling (Massaro et al. 2003; Saks et al. 2003) and sexual selection (Eley 1991; MacDougall and Montgomerie 2003; Massaro et al. 2003) in many species of birds and fish, may also affect the appearance of health in humans."
Race and physical attraction
Preferred and actual mate characteristics
The way individuals pair to produce reproductive units is a major factor determining evolution. This process is complex because it is determined not only by individual mating preferences, but also by numerous other factors such as competition between mates. Consequently, preferred and actual characteristics of mates obtained should differ, but this has rarely been addressed. We simultaneously measured mating preferences for stature, body mass, and body mass index, and recorded corresponding actual partner's characteristics for 116 human couples from France. Results show that preferred and actual partner's characteristics differ for male judges, but not for females. In addition, while the correlation between all preferred and actual partner's characteristics appeared to be weak for female judges, it was strong for males: while men prefer women slimmer than their actual partner, those who prefer the slimmest women also have partners who are slimmer than average. This study therefore suggests that the influences of preferences on pair formation can be sex-specific. It also illustrates that this process can lead to unexpected results on the real influences of mating preferences: traits considered as highly influencing attractiveness do not necessarily have a strong influence on the actual pairing, the reverse being also possible.
Mechanisms of sexual selection in humans
David A. Puts. Beauty and the beast: mechanisms of sexual selection in humans. Evolution and Human Behavior, Vol. 31, No. 3. (May 2010), pp. 157-175.A common refrain from certain non-whites and Southern Europeans is that if a Northern European man looks askance at the pursuit of Northern European women by non-Northern Europeans, it means he is "insecure" or afraid of competing with non-Northern European men. In reality, one competes by competing, and that starts (in a healthy society) with excluding men from other groups from access to women of one's own group.
Literature in evolutionary psychology suggests that mate choice has been the primary mechanism of sexual selection in humans, but this conclusion conforms neither to theoretical predictions nor available evidence. Contests override other mechanisms of sexual selection; that is, when individuals can exclude their competitors by force or threat of force, mate choice, sperm competition, and other mechanisms are impossible. Mates are easier to monopolize in two dimensional mating environments, such as land, than in three-dimensional environments, such as air, water, and trees. Thus, two-dimensional mating environments may tend to favor the evolution of contests. The two-dimensionality of the human mating environment, along with phylogeny, the spatial and temporal clustering of mates and competitors, and anatomical considerations, predict that contest competition should have been the primary mechanism of sexual selection in men. A functional analysis supports this prediction. Men's traits are better designed for contest competition than for other sexual selection mechanisms; size, muscularity, strength, aggression, and the manufacture and use of weapons probably helped ancestral males win contests directly, and deep voices and facial hair signal dominance more effectively than they increase attractiveness. However, male monopolization of females was imperfect, and female mate choice, sperm competition, and sexual coercion also likely shaped men's traits. In contrast, male mate choice was probably central in women's mating competition because ancestral females could not constrain the choices of larger and more aggressive males through force, and attractive women could obtain greater male investment. Neotenous female features and body fat deposition on the breasts and hips appear to have been shaped by male mate choice.
3.2.2.1. Male coalitions and between-group competition.
The tendency of males to form alliances may have evolved in the common ancestor of humans and our closest living relatives, Pan, as a means of cooperative female capture and defense (Fig. 3), although coalitions may also have evolved independently in these lineages for this purpose (Geary & Flinn, 2001; Wrangham, 1999). Male coalitions are rare among primates but common in humans and Pan, especially common chimpanzees (P. troglodytes), and are strengthened by kinship (Nishida & Hiraiwa- Hasegawa, 1987). The capture of women was a primary objective of early warfare (Darwin, 1871; Hrdy, 1997; Lerner, 1986; Spencer, 1885), and among foragers, groups of men commonly raid other villages and abscond with women (e.g., Chagnon, 1988). Such raids may also function in mate defense by deterring future attacks. These behaviors would tend to favor not only aggression and physical prowess, but also social intelligence for negotiating alliances (e.g., Alexander, 1989; Geary & Flinn, 2002; Mueller & Mazur, 1996; Wrangham, 1999).
Sheldon's t component
Assortative mating by genetic ancestry in admixed populations
within Mexican populations, people tended to pick partners with similar proportions of Native American and European ancestry, while in Puerto Rican populations couples had paired up based on their shared balance of European and African ancestry. [. . .] presumably people had cued into subtle variations in appearance, behaviour and even odour.The paper ("Ancestry-related assortative mating in latino populations") is open access.
"I think it's fascinating," says Burchard. "People are sizing up their partners, maybe in subconscious ways."